
ESCAN 2026
Thank you to the ESCAN board, organisers and staff, and to all the wonderful people I met, for the fantastic interest in my work and the thought-provoking talks.
Abstract
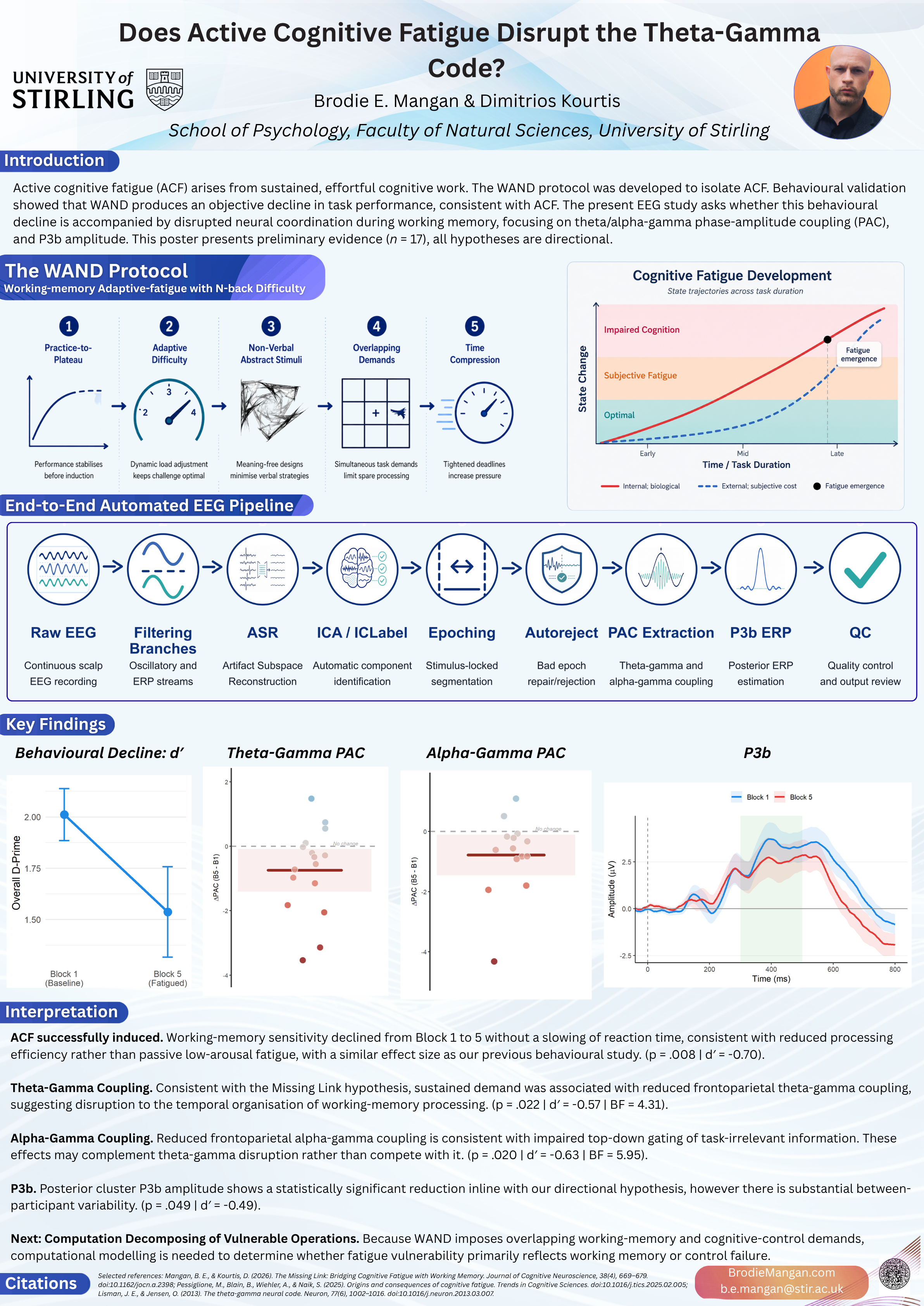
Cognitive fatigue is widely studied, yet few paradigms distinguish genuine active cognitive fatigue (ACF) from passive disengagement or task-learning artefacts. We previously validated the WAND (Working-memory Adaptive-fatigue with N-back Difficulty) protocol, demonstrating a robust decline in working-memory sensitivity (d′) with a medium-to-large effect size (Cohen’s d = −0.71), accompanied by response speeding rather than the slowing associated with passive fatigue (Mangan et al., preprint). However, the neural mechanisms through which ACF disrupts online information processing remain unspecified.
Here we test the Missing Link Hypothesis (Mangan & Kourtis, 2026): that ACF compromises the cross-frequency oscillatory dynamics that coordinate working memory function. Using EEG recorded during the WAND induction, we examine three complementary neural markers: (1) frontoparietal theta–gamma phase–amplitude coupling (TGC), associated with both the temporal organisation of working memory content and the cognitive control operations that regulate it; (2) frontoparietal alpha–gamma coupling (AGC), linked to top-down gating that suppresses task-irrelevant processing; and (3) posterior P3b amplitude, a broadly observed marker of task-directed attentional processing whose amplitude varies with stimulus relevance and available cognitive resources. Because the N-back task jointly engages working memory and cognitive control, disruption of these markers under ACF would indicate that the oscillatory architecture supporting sustained demanding performance is compromised, though it cannot alone resolve which function is primarily affected.
Supplementary Material
The Missing Link: From Cost Models to Mechanistic Targets
Recent fatigue models have become increasingly biologically grounded, explaining why sustained effort becomes harder to maintain through metabolic changes in control-relevant prefrontal regions (Holroyd, 2025; Pessiglione et al., 2025; Wiehler et al., 2022). However, these cost-of-control accounts do not specify how rising control costs translate into concrete failures of online information processing. Working memory is the natural place to look for that missing mechanism: it is the system through which currently relevant representations are maintained, prioritised, refreshed, and updated in the service of behaviour, making it a plausible interface between rising control costs and observable performance decline (D’Esposito & Postle, 2015; Miller et al., 2018).
Cross-Frequency Dynamics as Candidate Mechanisms
Working memory operates through coordinated oscillatory dynamics rather than a single static store. Two cross-frequency coupling mechanisms are of particular interest:
- Theta–Gamma Coupling (TGC): Theta–gamma coupling supports both the temporal organisation of working memory content (Lisman & Jensen, 2013) and the frontoparietal cognitive control operations that coordinate it (Daume et al., 2024; Friedrich et al., 2026; Miller et al., 2018). TGC is reduced in Alzheimer’s disease and mild cognitive impairment, where it is the strongest predictor of 2-back performance (Goodman et al., 2018), predicts individual differences in WM capacity in psychosis (Papaioannou et al., 2025), and serves as a functional marker of WM coordination difficulty in individuals with elevated autistic traits (Friedrich et al., 2026). In the N-back task, these functions are inseparable: the coupling may index the organisation of maintained items, the control operations that regulate them, or both.
- Alpha–Gamma Coupling (AGC): Alpha-phase modulation of gamma amplitude supports top-down gating, suppressing task-irrelevant processing to protect representational fidelity (Bonnefond & Jensen, 2015, 2025; Miller et al., 2018). Disruption of this mechanism should permit task-irrelevant information through, reducing the fidelity of currently active representations.
Importantly, even outside the fatigue context, there is substantial variability in cross-frequency coordination across individuals and performance states. Despite this variability, there is growing evidence that stimulating similar oscillatory markers can improve cognitive performance, including working memory. Cross-frequency stimulation targeting prefrontal theta–gamma synchronisation has enhanced spatial working memory (Alekseichuk et al., 2016), and individualised cross-frequency tACS targeting theta–gamma coupling has selectively modulated cognitive control performance and increased the targeted endogenous coupling pattern (Riddle et al., 2021). A recent meta-analysis confirms that active tACS produces reliable improvements in WM performance relative to sham (Nissim et al., 2023). If fatigue weakens the same frequency-specific dynamics that support performance and can be modulated by stimulation, then cross-frequency markers may provide candidate targets for future fatigue countermeasures.
Why Standard EEG Markers Are Insufficient
The fatigue literature has largely focused on broad spectral changes such as increased theta or alpha power (Tran et al., 2020). These markers index that fatigue is developing, but they are vague: they do not specify how cross-frequency neural coordination is affected. The ambiguity is consequential: an increase in frontal theta power, for example, could reflect a sleep-onset slowing signal, a compensatory upregulation of cognitive control to maintain performance, or a genuine fatigue-related disruption of processing efficiency. Without examining how these rhythms coordinate with one another, the same spectral observation can support contradictory interpretations. The present study bridges this gap by asking whether fatigue-related behavioural decline is accompanied by disruption in cross-frequency dynamics during a task that places sustained demand on both working memory function and cognitive control.
Next Steps
Establishing that ACF disrupts cross-frequency coordination addresses the mechanistic gap, but it opens a further question: which working memory and cognitive control operations are most vulnerable? Because the N-back task engages both simultaneously, future work will need to dissociate these functions using tasks that weight them differently, in order to identify which specific operation becomes vulnerable under fatigue and whether its disruption can be linked to a distinct neural signature.
Key References
- Alekseichuk, I., Turi, Z., Amador de Lara, G., Antal, A., & Paulus, W. (2016). Spatial working memory in humans depends on theta and high gamma synchronization in the prefrontal cortex. Current Biology, 26(12), 1513–1521. https://doi.org/10.1016/j.cub.2016.04.035
- Bonnefond, M., & Jensen, O. (2015). Gamma activity coupled to alpha phase as a mechanism for top-down controlled gating. PLoS ONE, 10(6), e0128667. https://doi.org/10.1371/journal.pone.0128667
- Daume, J., Kamiński, J., Schjetnan, A. G. P., et al. (2024). Control of working memory by phase–amplitude coupling of human hippocampal neurons. Nature, 629, 393–401. https://doi.org/10.1038/s41586-024-07309-z
- D’Esposito, M., & Postle, B. R. (2015). The cognitive neuroscience of working memory. Annual Review of Psychology, 66, 115–142. https://doi.org/10.1146/annurev-psych-010814-015031
- Goodman, M. S., et al. (2018). Theta-gamma coupling and working memory in Alzheimer’s dementia and mild cognitive impairment. Frontiers in Aging Neuroscience, 10, 101. https://doi.org/10.3389/fnagi.2018.00101
- Lisman, J. E., & Jensen, O. (2013). The theta-gamma neural code. Neuron, 77(6), 1002–1016. https://doi.org/10.1016/j.neuron.2013.03.007
- Bonnefond, M., & Jensen, O. (2025). The role of alpha oscillations in resisting distraction. Trends in Cognitive Sciences, 29(4). https://doi.org/10.1016/j.tics.2025.02.003
- Mangan, B. E., & Kourtis, D. (2026). The missing link: Bridging cognitive fatigue with working memory. Journal of Cognitive Neuroscience, 38(4), 669–679. https://doi.org/10.1162/JOCN.a.2398
- Miller, E. K., Lundqvist, M., & Bastos, A. M. (2018). Working memory 2.0. Neuron, 100(2), 463–475. https://doi.org/10.1016/j.neuron.2018.09.023
- Pessiglione, M., Blain, B., Wiehler, A., & Naik, S. (2025). Origins and consequences of cognitive fatigue. Trends in Cognitive Sciences, 29(8), 730–749. https://doi.org/10.1016/j.tics.2025.02.005
- Riddle, J., et al. (2021). Brain-based individualization of frequency-coupled transcranial alternating current stimulation for cognitive control. Journal of Cognitive Neuroscience, 33(8), 1523–1537. https://doi.org/10.1162/jocn_a_01732
- Tran, Y., Craig, A., Craig, R., Chai, R., & Nguyen, H. (2020). The influence of mental fatigue on brain activity: Evidence from a systematic review with meta-analyses. Psychophysiology, 57(5), e13554. https://doi.org/10.1111/psyp.13554
- Friedrich, E. V. C., Hilla, Y., Sterner, E. F., Ostermeier, S. S., Behnke, L., & Sauseng, P. (2026). Theta-gamma phase amplitude coupling serves as a marker of social cognition and visual working memory deficits in individuals with elevated autistic traits. Communications Psychology, 4, 25. https://doi.org/10.1038/s44271-025-00392-6